DNA双螺旋结构发现70周年:自我复制的生命演化史!
Posted turingbooks
tags:
篇首语:本文由小常识网(cha138.com)小编为大家整理,主要介绍了DNA双螺旋结构发现70周年:自我复制的生命演化史!相关的知识,希望对你有一定的参考价值。


2023年是DNA双螺旋结构发现七十周年。1953年2月28日,沃森和克里克根据女科学家弗兰克林拍摄的X射线衍射照片,成功推断出遗传物质DNA的三维结构模型。
砖头瓦块已经齐备,建筑师也已经充满能量,随时准备不辞辛苦地修建起辉煌壮丽的生命大厦,但是对于任何一种能够抗拒亿万年风霜摧折、在地球上生存和繁盛的生命来说,仅仅有这些还远远不够。
原因很容易理解:这座大厦太脆弱了。
万一一场地震或者火灾毁掉了唯一的大厦怎么办?万一大厦的基础被蝼蚁松动,或者一场台风卷走了大厦的顶层呢?要知道,概率再小的意外,放在几十亿年的时间尺度中,都会变得实实在在起来。也就是说,仅仅由能量驱动建立起来的生命大厦,即便真的在远古地球上出现过,恐怕也早在漫长的时光里毁于意外事故了,那么今天的地球人类估计就没有缘分看到这样的生命了。
更要命的原因还不在这里,而在于地球环境不是永恒不变的!在我们每个人几十年的生命中,我们也许可以安心期待日复一日的日升日落、年复一年的春夏秋冬。当然,这一切还得期待全球气候变化不会带来灾难性的后果。但是如果把时间尺度放大到生命演化的尺度——几千万年到几十亿年,我们就会发现,地球环境的变化剧烈得远非“沧海桑田”几个字所能概括。
举一个我们可能会熟视无睹的例子吧:氧气。人类生存需要氧气,这是因为在人体中,能量货币 ATP 的生产过程严重依赖氧气。这一点我们已经讨论过,在氧气缺乏的环境下(例如肌肉持续收缩时),一个葡萄糖分子分解成两个乳酸分子,仅仅能产生两个 ATP 分子。而在氧气的帮助下,葡萄糖分子可以彻底分解为二氧化碳和水。这个过程中所释放的能量,通过驱动细胞内的微型水电站——ATP 合成酶,可以制造出多达 28~38 个 ATP 分子。因此,如果没有氧气,人体将无法进行永不停歇的生命活动——从心脏跳动、游泳跑步,到思维和语言。实际上,人体对氧气浓度的适应区间是非常狭窄的。在海拔四五千米的青藏高原,氧气浓度下降到 10% 多一点,人体就会出现缺氧的症状。反过来,如果吸入的氧气浓度过大,人体就会出现所谓的“氧中毒”现象,神经系统、肺和眼球都会受到严重损伤。同样的例子还有温度。人体适宜的环境温度在 25 摄氏度上下浮动。如果人体长期处于 40 摄氏度以上的环境中,很容易引起中暑死亡;处于低温环境下也不行,人在 5 摄氏度的海水里只能活个把小时。
特别是,如果考虑更长的时间尺度的话,我们会发现,在亿万年的生物演化历史上,能够满足人体生存环境要求的时间段实在是太狭窄了!在过去六亿年的时间里,大气氧含量可能在 5% 到 35% 之间反复剧烈波动,平均气温的变化范围是 10~40 摄氏度(作为参照,如今地球的平均气温大约是 15 摄氏度)。请注意,这里我们仅仅考虑了过去六亿年,并且只考虑了氧气浓度和气温两个环境指标。如果把时间尺度扩大到整个地球生命史,再考虑到太阳光强度、昼夜长短、大气组成、土壤的化学成分、食物和天敌等复杂的环境因素,就会得到一个不言而喻的结论——在翻脸无情的地球母亲的怀抱里,没有哪个生命可以做到永远左右逢源。
如果生命真的是一座大厦,那么不管修建的时候用了多么坚固耐用的砖瓦,在建成时是多么辉煌壮丽,考虑到它一会儿会在阳光下暴晒,一会儿淹没在倾盆大雨中,一会儿又要被坚冰覆盖,时而被蝼蚁侵蚀,时而受猛兽冲撞,时而遭受流星雨和地震的摧折,它绝不可能永远基业长青。
一个显而易见的悖论出现了。诞生于远古地球的始祖生命——那些由能量这个天才建筑师建立的生命大厦——是如何逃开了无可避免的意外事故和难以抗拒的沧桑巨变,绵延不绝一直到今天的?
答案其实很简单:自我复制。
自我复制是地球生命基业长青的基础——以自身为样本,不停地制造出和自己相似但又不完全一样的子孙后代。
后代越来越多,就保证了即便其中一些因为意外事故——不管是台风、地震还是蝼蚁——死去,还有足够的个体能存活下来延续香火。
而更重要的是,(不够精确的)自我复制为生命现象引入了变化。这种变化大多数时候难以察觉,比如生命大厦悄悄更换了天井的绿植或是大堂的灯饰。但有些时候也可以惊天动地,整座大厦的楼高、外饰面乃至主干结构都焕然一新。但是无论如何,在自我复制过程中产生的变化,总是快过地球环境动辄以千万年计数的变化。也正因为这样,地球上的生命来了又走,样貌也千变万化——科学家的估计是,在这颗星球上,可能已经有超过 50 亿个物种诞生、繁盛,然后静悄悄地死去——但是生命现象本身却顽强地走过了 40 亿年的风霜雨雪。
当然,在自我复制中出现的这些不怎么引人瞩目的细微变化本身谈不上什么对错,也没有什么方向性可言。不够精确的自我复制,其实是提供了大量在地球环境中“试错”的生物样品。谁能活下来,谁能继续完成新一轮自我复制,谁就是胜利者。是地球环境的缓慢变迁决定了不同时刻的胜利者,也因此最终塑造了生物演化的路径。
比如,我们刚才说到从六亿年前到今天,大气中氧气的含量始终在上下波动。但是如果时间尺度放得更宽,我们会发现氧气甚至压根儿就不是地球上从来就有的大气成分。在 46 亿年前地球形成的时候,大气的主要成分是二氧化碳、氮气、二氧化硫和硫化氢。直到差不多 25 亿年前,第一批能够利用阳光的细菌出现在原始海洋中,利用太阳光的能量分解大气中的二氧化碳,并以其中的碳原子为食,这才制造出了氧气。对于今天的地球生命无比重要的氧气,其实在当时只是某些生命活动的副产品。更可怕的是,这种全新的化学物质还毒死了当时地球上几乎所有的生物!但是与此同时,灾难性的“大氧化”事件却为未来那些以氧气为生、更复杂多样的生命开启了繁盛的大门,受益者包括海藻、树木、鱼和人类。那些能够在无氧大气里生息繁盛的生命和那些在氧气中自在生活的生命,并无高下之别,仅仅是由于地球环境的变化让前者死去、后者存活罢了。
因此,自我复制的两个看起来似乎自相矛盾的特点保证了地球生命的永续。对自身的不断复制保证了生命不会因一场意外而彻底毁灭,而自我复制过程中出现的错误,则帮助生命适应了地球环境的变化。
那么,自我复制又是怎么发生的呢?

01
思想实验中的生命演化史
我们不妨先做一个思想实验,构造一个极端简化的生命,探讨一下生命自我复制的原理。
从最简单的情形开始,我们思想实验中的生命——就叫它生命 1.0 吧——只有一个蛋白质分子。从前面的故事里大家可以很容易想象,最有资格入选的蛋白质大概就是为生命制造能量货币的 ATP 合成酶了。这个古老的蛋白质分子尺寸很小,仅有个纳米那么大,却蜷曲折叠成一个复杂的、带有三个叶片和一个管道的三维结构,通过飞速旋转不停地生产 ATP。有了它,生命 1.0 就可以制造 ATP 分子,然后用 ATP 来驱动各种生命活动了。
但是生命 1.0 是难以实现自我复制的。从前一章中我们知道,ATP 合成酶有一个极端精巧和复杂的三维立体结构,每个维度上原子排列的精确度达到零点几纳米的水平。且不说想要分毫不差地复制一个这样的结构非常困难,即便是想要看得清楚一点都不容易。在今天人类的技术水平下,要看清楚 ATP 合成酶的每一个原子,需要动用最强大的 X 射线衍射仪和电子显微镜,而想要复制出这样一个结构,还是科幻想象的范畴。
我们大概可以说,要想一丝不苟地复制生命 1.0,可能需要一架比生命 1.0 体形更庞大、更加复杂和精密的机器才做得到。可是在刚刚出现生命 1.0 的远古地球上,又去哪里找这样的复杂机器呢?难以自我复制的生命 1.0 注定要孤独一生——而且它的一生一定非常短暂。
为了解决自我复制的技术困难,生命体显然需要一种方法,能更简单精确地记录和复制自身,不要让我们瞪大眼睛去记录和复制一个复杂三维结构的每一点空间信息。这样太烦琐,也太容易出错了。
于是,生命 2.0 应运而生。人体中的 ATP 合成酶是由五千多个氨基酸分子按照某种特定顺序串起来形成的蛋白质大分子。在三维空间中,这些氨基酸彼此吸引、排斥、碰撞、结合,形成了复杂、动态的三维结构。那么可想而知,只要我们能记录下这五千多个氨基酸分子的先后顺序,然后依样画葫芦地依照这个顺序去组装 ATP 合成酶分子就行了,它可以自己完成在三维空间的折叠扭曲。这样一来,三维空间的信息就被精简成了一维,只是一组顺序排列的氨基酸分子而已。
在今天的绝大多数地球生命中,三维到一维的信息简化是通过 DNA 分子实现的。DNA 分子的化学构成其实非常简单,就是由四种长相平凡的核苷酸分子环环相扣串起来的一条长长的链条。它的秘密隐藏在四种核苷酸分子的排列组合顺序中(见图 3-1)。在今天的地球生命体内,DNA 长链按照三个核苷酸的排列顺序决定一个氨基酸的原则,能够忠实记录任何蛋白质分子的氨基酸构成——当然也包括生命 1.0 中的 ATP 合成酶。
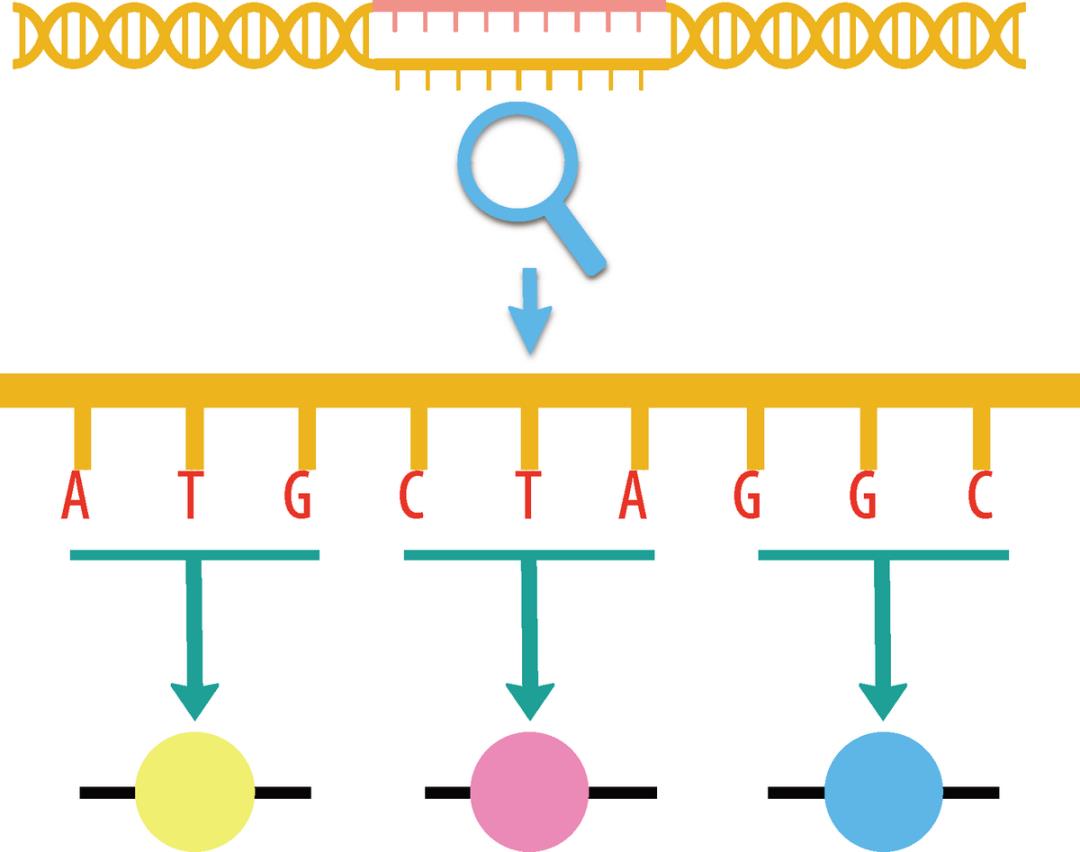
图 3-1 三个碱基密码子对应一个氨基酸。在今天绝大多数的地球生命中,DNA 链上的核苷酸分子会按照三三一组形成“密码子”,每组密码子对应一种氨基酸
这样一来,生命 2.0 在自我复制的时候,就不需要担心复杂的 ATP 合成酶蛋白无法精细描摹和复制了。它只需要依样画葫芦地复制一条 DNA 长链就行了,因为 DNA 长链本身的组合顺序就已经忠实记录了 ATP 合成酶的全部信息(见图 3-2)。而与此同时,我们可以想象,复制一条可以看成是一维的 DNA 长链,要比直接复制 ATP 合成酶的精细三维结构省心省力得多。

图 3-2 DNA 的复制模型。首先,一条 DNA 双螺旋链一分为二;随后,两条 DNA 单链分别作为模板,与新加入的碱基分子配对,形成两条新 DNA 双螺旋
在 20 世纪 50 年代,詹姆斯 • 沃森(James Watson)和弗朗西斯 • 克里克(Francis Crick)利用罗莎琳 • 富兰克林(Rosalind Franklin)获得的 X 射线衍射图谱,建立了 DNA 的双螺旋模型,并且几乎立刻猜测到了 DNA 是如何进行自我复制的。简单来说,DNA 复制遵循的是“半保留复制”的机制。就像蛋白质是由 20 种氨基酸砖块组合而成的,DNA 也有它独有的砖块:四种不同的核苷酸分子(可以简单地用 A、T、G、C 四个字母指代)。每条 DNA 链条都是由这四种砖块首尾相连组成的。特别重要的是,ATGC 四种分子能够两两配对形成紧密的连接:A 和 T,G 和 C。因此可以想象,一条顺序为 AATG 的 DNA 可以和一条 CATT 的 DNA 首尾相对地配对组合缠绕在一起。这样的配对方法特别适合 DNA 密码的自我复制:AATG 和 CATT 两条缠绕在一起的链条首先分离开来,两条单链再根据配对原则安装上全新的核苷酸分子,例如 AATG 对应 CATT,而 CATT 则装配了 AATG,由此一个 DNA 双螺旋就变成了完全一样的两个 DNA 双螺旋。特别值得指出的是,DNA 的复制过程异常精确,在人体细胞中,DNA 复制出错的概率仅有 10 的 9 次方分之一。这就从原理上保证了它可以作为生命遗传信息的可靠载体。
那么,简单而强大的生命 2.0 能否稳定地生存和繁衍呢?
很遗憾,还是不行。事实上它根本就不可能存在。DNA 是一种化学上非常稳定的分子——这并不奇怪,能被选中作为信息密码本的化学分子当然必须稳定和可靠,甚至是懒惰。也正因此,一根光秃秃的 DNA 长链根本就什么都不会干,它既不可能自我复制,也不可能制造出什么 ATP 合成酶来。就像一本写满了字母的密码本,要是没有人抄写,没有人解读,它自己什么也做不了。
好吧,不气馁的我们继续升级出了生命 3.0。这次,我们需要的东西就多了许多。除了负责制造能量货币的 ATP 合成酶之外,生命 3.0 还需要一大堆各种各样的蛋白质分子,来实现 DNA 分子的自我复制,利用 DNA 分子携带的信息制造各种新的蛋白质。
单单说 DNA 复制就已经非常复杂了。生命 3.0 需要蛋白质分子帮忙把高度折叠的 DNA 展开变成长链(就拿人的 DNA 来说,完全伸展开来长达数米,所以必须经过几轮折叠包装,才能塞进直径仅有几微米的细胞里,仅仅在使用时才部分展开),需要蛋白质分子在 DNA 长链上精确定位到底从哪里开始复制,需要蛋白质分子运送 DNA 复制所需的原材料(比如四种核苷酸分子),需要蛋白质分子填补复制当中的缺口,修正复制过程中出现的错误,还需要蛋白质分子将复制好的 DNA 长链重新折叠回去。
同时,生命 3.0 还需要一大堆蛋白质分子(见图 3-3),根据 DNA 密码本的信息制造 ATP 合成酶——有的负责读取 DNA 密码本的信息,有的负责搬运蛋白质的原料氨基酸,有的负责氨基酸的装配顺序,等等。当然了,再多的蛋白质分子也难不倒我们,我们可以利用在生命 2.0 中就确定的规则,把它们的信息也写入 DNA 长链中去,这样仍然是复制一条 DNA 长链,生命 3.0 就可以把所有蛋白质分子的信息都忠实地复制和传递下去了。
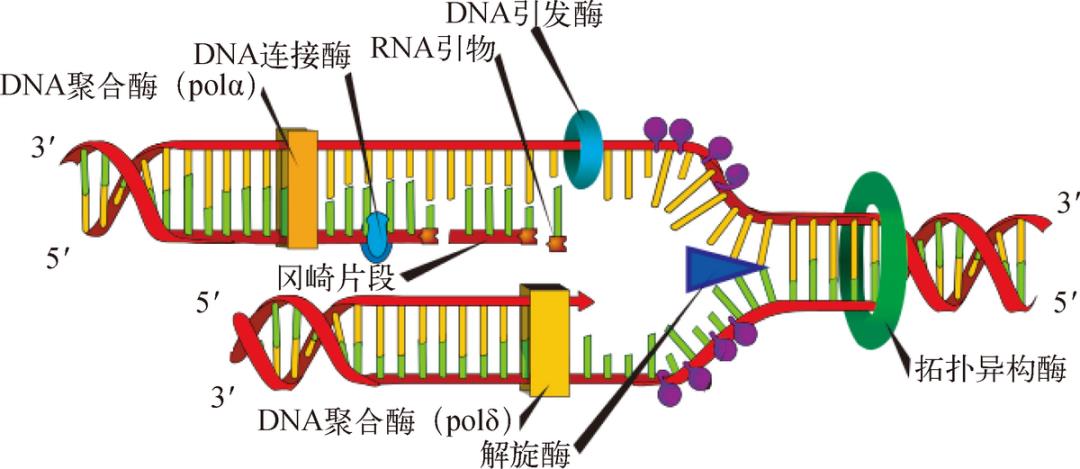
图 3-3 DNA 复制所需的蛋白质。图中仅仅呈现了极小的一部分,但我们已经可以看到,DNA 复制是一个需要大量蛋白质帮手参与的精细过程。DNA 双链首先会在拓扑异构酶和解旋酶的帮助下分解成单链。随后,不同的 DNA 聚合酶分别负责两个方向的 DNA 复制,复制完成的短片段还要在连接酶的帮助下连成长片段
生命 3.0 的命运如何?
我们已经很接近成功了,但是还差重要的一点点。
根据前面的描述,大家就能猜想到,蛋白质分子和 DNA 长链,对于生命的生存和复制来说,是相辅相成缺一不可的。前者的制造依赖于后者保存的信息,而后者也依赖前者完成自我复制,因此空间上它们必须离得足够近才行。我们必须想出一个办法,把这些东西统统聚拢到一起,保护起来。否则,蛋白质和 DNA 都很容易在自然环境中扩散得无影无踪,谁也找不到谁。
解决方案倒也不难想,用一张致密的网把所有这些林林总总的蛋白质和 DNA 都给包裹起来就行了。在今天的地球生命中也有这张网,名字叫作细胞膜,是一层仅有几纳米厚度、由脂肪分子构成的薄膜。这层薄膜紧紧地包裹住了蛋白质和 DNA,形成了一个细胞,把它们和危险的自然环境隔绝开来。在今天的地球生命里,除了少数例外(比如病毒),绝大多数生命都是由一个或者多个细胞组成的。
这次,就叫它生命 4.0 吧。
目前,生命 4.0 已经有点极简版地球生命的样子了。我们权且相信它能够在地球上生存下来,因为它能够不断地从环境中攫取能量供给生命活动,也能不停地自我复制对抗衰退和死亡。实际上,今天的地球生命尽管比我们思想实验中的生命 4.0 要复杂得多,但是从基本原理上看,确实相差无几。
但是新的问题来了:这个看起来靠谱的生命 4.0,真的有可能在 40 亿年前魔法般地出现在地球上吗?换句话说,生命 4.0 的构想固然有它的内在逻辑,但它真的有可能模拟了地球生命的最初起源吗?
很遗憾,答案是不可能。或者,至少看起来非常不可能。
其中的麻烦有点像“鸡生蛋还是蛋生鸡”的问题。蛋白质的全部信息都存储在 DNA 密码本中,依靠 DNA 密码本中忠实记录的信息,我们能够制造出各种各样的蛋白质分子。因此,让我们权且假定 DNA 是“鸡”,蛋白质是它下的“蛋”。
但是,一条孤零零的 DNA 长链是没有办法干任何事情的,它需要各种蛋白质分子的帮忙,才能实现自我复制,需要依赖蛋白质的帮忙才能制造出新的蛋白质。如果没有提前准备好蛋白质“蛋”,DNA“鸡”根本没法继续生“蛋”!
换句话说,我们设计的生命 4.0 想要自发出现,我们得不断祈求大自然同时造就信息互相匹配的“鸡”和“蛋”。而且,“鸡”和“蛋”还必须几乎同时出现,距离无比接近,才有可能配合起来造就生命。要是在一阵电闪雷鸣中,一只 DNA“鸡”被创造了出来,但是它附近却没有那只冥冥中注定属于它的蛋白质“蛋”,那么这只 DNA“鸡”只能沉默着走向分解破碎,因为它自己什么也做不了。而反过来,要是那个蛋白质“蛋”率先在海底的热泉口奇迹现身,甚至还能工作一下或生存一会儿,但是因为没有 DNA“鸡”帮它保留和传递信息,“蛋”必然也会快速走向衰退和死亡。
那怎么办?地球生命对这个问题的回答是非常耐人寻味的:既不是先有蛋,也不是先有鸡。事实上,很可能在生命刚刚出现的时候,鸡和蛋都还没有踪影呢。

02
不是鸡不是蛋,既是鸡又是蛋
让我们再回顾一下生命 4.0 的基本设计原则吧。DNA 负责记录蛋白质分子的氨基酸排列信息,以 DNA 序列为模板可以制造出各式各样的蛋白质分子。而反过来,蛋白质分子除了制造能量,还可以帮助 DNA 实现自我复制。这好像是个挺简单的二元系统,是不是?
在自然界,简单往往意味着高效、节约和更容易自发出现。但是很让人意外的是,地球生命不约而同地选择了一种更复杂、相对也更容易出错和更浪费的办法:在 DNA 和蛋白质的二元化结构之间,平白无故地多了第三者:RNA(ribonucleic acid,核糖核酸)。
RNA 是一种长相酷似 DNA 的化学物质,两者的唯一区别就是化学骨架上的一个氧原子。对于我们的生命 4.0 系统来说,RNA 像二郎神的第三只眼睛一样,显得非常怪异和多余。当生命开始活动的时候,DNA 密码本的信息首先被忠实地誊抄到 RNA 分子上,然后 RNA 分子再去指导蛋白质的装配。放眼望去,加上 RNA 的生命——就叫它生命 5.0 好了——实在是看不出有什么优势来。打个比方,原本在车间里,一个经理直接指导工人干活就挺好的,命令传达简单快捷还不容易出错。现在非要给经理配一个主管,每一道命令都必须由经理告诉主管,主管再告诉工人。直觉告诉我们,这样的系统一定存在命令走样变形、人际关系复杂多变等问题,更不要说还得多付这个主管的工资了!
然而,这套叠床架屋的所谓“中心法则”(见图 3-4)几乎成了所有地球生命运转的核心,既保证了遗传信息的世代流传,也保证了每一代生命体实现自身的生命机能。这种巨大的反差驱使人们从反方向思考,也许 DNA → RNA →蛋白质的系统有极其深远但仍不为人所知的意义,以至于这个看起来如此多余、低效和浪费的系统能够挺过严酷多变的地球环境和物种竞争,保留在绝大多数地球生命的身体里。
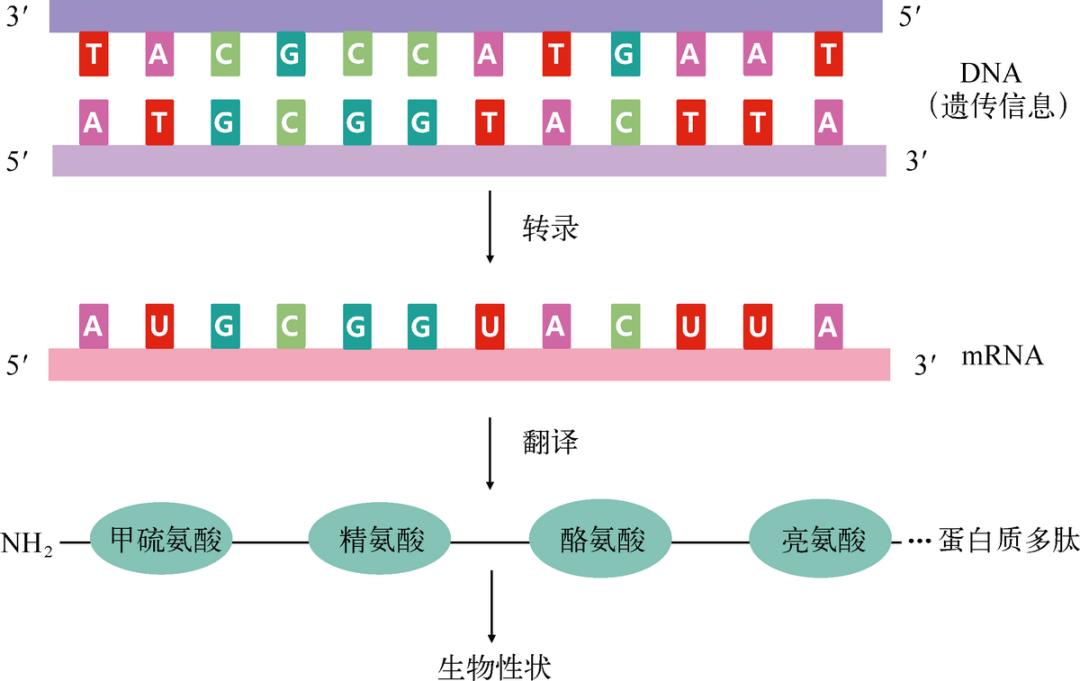
图 3-4 生命的“中心法则”。依据中心法则,DNA 的自我复制保证了遗传信息的传递和生命的生生不息,DNA 也通过指导蛋白质合成决定了生命活动的形态。RNA 的产生则是其中的一个中间步骤,RNA 一方面忠实抄写了 DNA 的密码信息,另一方面直接指导了蛋白质的制造。值得指出的是,地球生命中也有不少中心法则之外的生命。比如某些病毒并没有 DNA,而是直接利用 RNA 来存储遗传信息并指导蛋白质合成(例如流感和丙肝病毒)。也有一些病毒虽然使用 DNA,但是和图中不同,它们仅有一条单链 DNA,只在启动自我复制的时候才变成双螺旋
这样做的意义是什么呢?
事实上,早在 20 世纪中叶,当 DNA → RNA →蛋白质这套遗传信息传递的所谓“中心法则”刚刚被提出的时候,就已经有人问这样的问题了。例如 1968 年,DNA 双螺旋的发现者之一克里克就在一篇文章中大胆地猜测,也许看起来多余的 RNA 才是最早的生命形态。他甚至说:“我们也不是不能想象,原始生命根本没有蛋白质,而是完全由 RNA 组成的。”但是猜想毕竟只是猜想,看似无用的 RNA 反而可能是最早的生命,重要的 DNA“鸡”和蛋白质“蛋”反而仅仅是 RNA 的后代和附属品。这样的想法可以引发很多哲学上有趣的思考,但是很少有人期待真的在自然界或者实验室里验证它。
直到 1978 年,30 岁的生物化学家汤姆 • 切赫(Tom Cech)来到美丽的山城——美国科罗拉多州的邦德建立了自己的实验室。
他的研究兴趣和我们讲过的中心法则有密切的关系。我们知道,在遗传信息的流动中,RNA 是承接在 DNA 和蛋白质之间的分子。它誊抄了 DNA 密码本的信息,然后再以自身为蓝图,指导蛋白质的装配。不过早在 20 世纪 60 年代,人们就已经发现,RNA 密码本其实并不是一字不差地誊抄了 DNA 密码本的信息,例如 DNA 密码本中往往会写着大段大段看起来没有什么特别用处的“废物”字母(它们的学名叫作“内含子”)。在抄写 RNA 密码本的时候,生物会首先老老实实地誊抄这些废物字母,之后再将它们整页撕去,整理出更精简更经济的一本密码本。
切赫当时的兴趣就是研究这种被叫作“RNA 剪接”——也就是如何撕去密码本中间多余的纸张——的现象。他使用的研究对象是嗜热四膜虫 1(tetrahymena thermophila),这是一种分布广泛的淡水单细胞生物,很容易大量培养,并且个头很大(直径有 30~50 微米),很方便进行各种显微操作。而研究 RNA 剪接也是分子生物学黄金年代里热门的话题之一,毕竟它关系到遗传信息如何最终决定了生物体五花八门的生物活动和性状。
1嗜热四膜虫这种看起来不起眼的单细胞生物孕育了 20 世纪的许多伟大发现。除了下文会讲到的核酶和 RNA 世界,还有对于衰老异常重要的端粒和端粒酶(2009 年诺贝尔生理学或医学奖),以及蛋白质的翻译后修饰等。
一开始,切赫的目标是很明确的。他已经知道,在四膜虫体内的 RNA 分子中段,有一截序列是没有什么用的。这段被称为“中间序列”的无用信息,在 RNA 刚刚制造出来之后很快就会被从中间剪切掉。而这个过程是怎么发生的呢?切赫希望利用四膜虫这个非常简单的系统来好好研究研究。他的猜测也很自然:肯定有那么一种未知的蛋白质,能够准确地识别这段 RNA 中间序列的两端,然后“咔嚓”一刀切断 RNA 长链,再把两头缝合起来,RNA 剪接就完成了。
为了找出这个未知的蛋白,切赫的实验室使用了最经典的化学提纯方法。他们先准备了一批尚未切割的完整 RNA 分子,再加入从四膜虫细胞中提取出来的蛋白质混合物“汤”。那么显然,RNA 分子应该会被切断和缝合,从而完成密码本的精简步骤。他们的计划是,把蛋白质“汤”一步一步地分离、提纯,排除掉那些对 RNA 剪接没有影响的蛋白质,那么最终留下的应该就是他们要找的那个负责剪接 RNA 的蛋白质了。
但是,他们的尝试刚一开始就差点胎死腹中。因为切赫发现,RNA 分子加上蛋白质“汤”确实会很顺利地启动剪接。但是即便什么蛋白质都不加,RNA 分子也同样出现了剪接!
任何一个受过起码的科学训练的人都明白,这个现象是多么令人沮丧。什么都不加的 RNA 分子也能被剪接,看起来只有两个可能性:第一,切赫他们制备的 RNA 已经被污染了,里面混入了能够切割 RNA 的蛋白质,因此不管加不加东西,RNA 分子都被剪接了;第二,切赫他们看到的这个现象压根儿就不是 RNA 剪接,而是一种不知道是什么的实验错误,因此加不加其他蛋白质,他们看到的都不是剪接。不管是哪种解释,眼看着这个实验就做不下去了。
于是,切赫他们尝试了各种各样的办法来改进实验。他们首先假定自己的纯化功夫确实不到位,RNA 被污染了,因此想要从里面找出那种被“污染”的蛋白是什么,没成功;后来他们往纯化出的 RNA 分子里加上各种各样破坏蛋白质活性的物质,试图停止 RNA 的剪接,发现也不成功;他们甚至还做了更精细的化学实验,来研究 RNA 到底是怎么被剪接的、发生了什么化学修饰……
终于,到了 1982 年,切赫他们干脆放弃了对 RNA 分子各种徒劳的提纯,直接在试管里合成了一个新的 RNA 分子。然后,利用这条理论上就不可能存在污染的纯净 RNA,他们终于可以明白无误地确认,这条 RNA 在什么外来蛋白质都没有的条件下,仍然固执地实现了自我剪接,把那段没用的中间序列切割了出来。
事情已经无可置疑。根本不存在那种看不见摸不着又总是顽固地剪接 RNA 的蛋白质,RNA 可以自己剪断和粘连自己!
说得更酷一点,原本大家觉得多余和浪费的 RNA 分子,居然可以身兼 DNA 和蛋白质的双重功能:它显然可以和 DNA 一样存储信息,同时也可以像蛋白质一样催化生物化学反应——在切赫的例子里,这个化学反应就是对自身进行切割和缝合。切赫给他们找到的这种新物质命名为“核酶”(ribozyme,兼具核酸和酶的功能之意,见图 3-5),而科学界也闪电般地以 1989 年诺贝尔化学奖回报了这个注定要名垂青史的伟大发现。
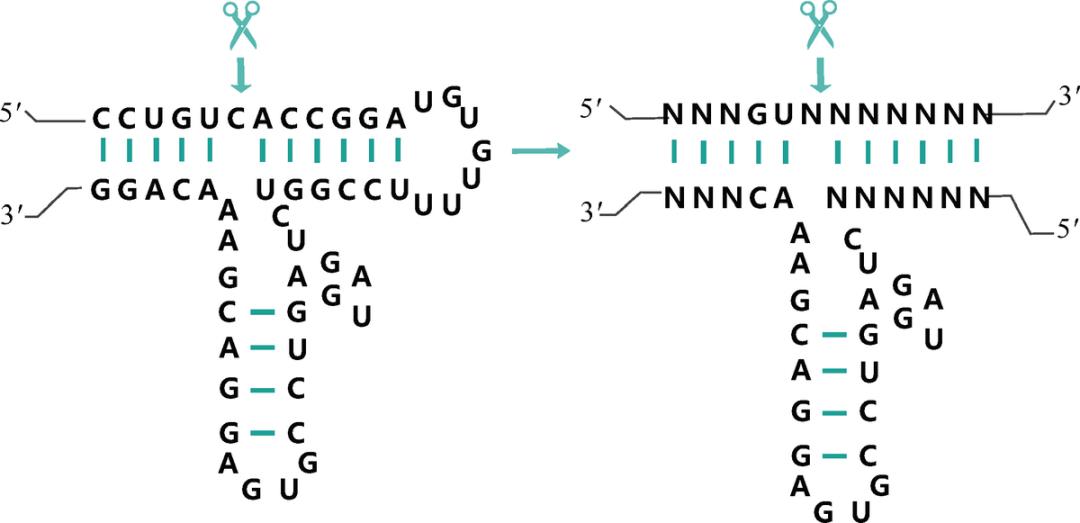
图 3-5 “锤头”核酶(hammerhead ribozyme),广泛存在于从细菌到人体的多种生物中。一条 RNA 链能内部折叠配对,形成一种状似发卡的结构,并能在特定位置实现自我剪切
当然无论如何,剪接这件事本身看起来其实不过是一种并不那么复杂的生物化学反应。无非是“咔嚓”一刀,然后缝好,在地球生命的体内只能算是一种平淡无奇的现象。但是这个发现仍然具有极其深远的意义,既然在一种单细胞浮游生物体内确实存在着一种分子,它既可以作为密码本记录生命体的遗传信息,又可以作为分子机器驱动一种简单的生命活动过程,那么举一反三,就会想到,也许曾经存在一大类逻辑类似的核酶分子,它们既能够记录五花八门的遗传信息,又能够实现形形色色的生命机能,一身担起 DNA 和蛋白质的使命。
而只要稍加推广,我们就会发现,核酶的概念似乎可以用来解释生命起源!不是说 DNA 和蛋白质先有鸡还是先有蛋这个问题无法解决吗?核酶这种奇怪的东西,至少理论上可以既是鸡又是蛋!只要想象一个这样的 RNA 分子,它自身携带遗传信息,同时又能催化自身的复制(相比剪接,这当然是一种复杂得多的生物化学反应),那不就可以实现遗传信息的自我复制和万代永续了吗?什么 DNA,什么蛋白质,对于伟大的生命起源来说,不过是事后锦上添花的点缀而已!

04
RNA世界
不得不承认,这个思路的脑洞还是开得很大的。要知道,虽然切赫发现的核酶确实实现了一点替代蛋白质的功能,但这个功能还是非常简单的,只是给 RNA 做个砍头接脚的外科手术而已。而如果真要设想一种核酶能够实现自我复制的功能,它必须能够以自身为样本,把一个接一个的核苷酸按照顺序精确地组装出一条全新的 RNA 链条来。这个难度比起 RNA 剪接,简直是汽车流水线和榔头剪刀的差别。
不过没过多久,大家在研究细胞内的蛋白质生产过程的时候,就意识到 RNA 的能力远超人们的想象。我们知道,蛋白质分子的生产是以 RNA 分子为模板,严格按照三个核苷酸分子对应一个氨基酸分子的逻辑,逐渐组装出一条蛋白质长链的过程。这个过程是在一个名叫“核糖体”(模型见图 3-6)的车间里进行的。而从 20 世纪 80 年代开始,人们在研究核糖体的时候逐渐意识到,这个令人眼花缭乱的复杂分子机器,居然是以 RNA 为主体形成的!在细菌中,核糖体车间的工作人员包括 50 多个蛋白质,以及三条分别长达 2900、1600 和 120 个核苷酸分子的 RNA 链。这些 RNA 链条上的关键岗位对于决定蛋白质生产的速度和精度至关重要。

图 3-6 核糖体模型
既然连蛋白质生产这么复杂的工作 RNA 都可以胜任,那还有什么理由说,在生命诞生之初,RNA 分子就一定不能做到自我复制呢?
就是在这样脑洞大开的思路指引下,全世界展开了发现、改造和设计核酶的竞赛。我们当然没办法看到地球生命演化历史上第一个自我复制的核酶到底是什么样子的,但是如果人类科学家能在实验室里人工制造出一个能够自我复制的核酶,我们就有理由相信,具备同样能力的分子在远古地球上出现,并不是什么天方夜谭。
2001 年,美国麻省理工学院的科学家成功“制造”出了一种叫作 R18 的、具有部分自我复制功能的核酶分子(见图 3-7),第一次证明核酶确实不光能当榔头剪刀,还真的可以装配汽车!当然,R18 的功能还远不能和我们假想中的那个既能当鸡又能当蛋的祖先 RNA 相比,R18 仅仅能够复制自身不到 10% 的序列,而我们的祖先可一定需要 100% 复制自身的能力。但这毕竟是一个概念上的巨大突破。要知道,既然人类科学家可以在短短几年内设计出一个具备初步复制能力的核酶,那么我们就没有理由怀疑,无比浩瀚的地球原始海洋在几亿年的时间里会孕育出一个真正的祖先核酶。
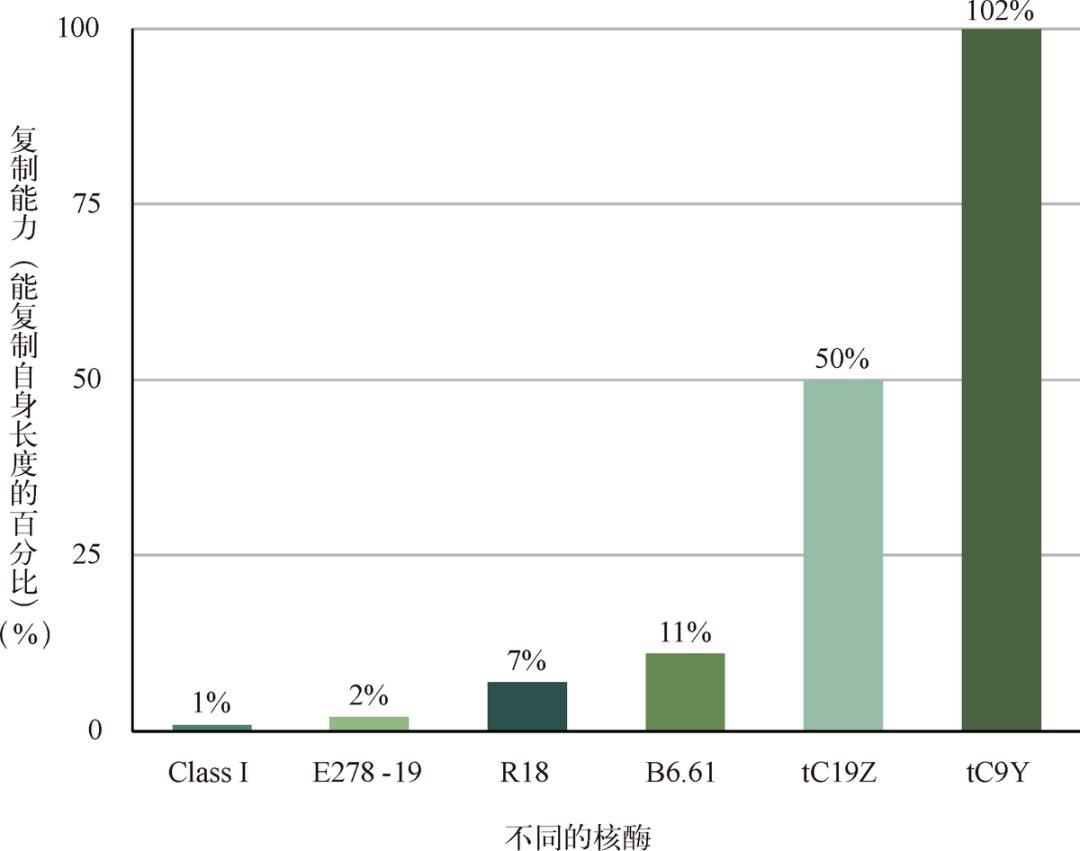
图 3-7 人工设计核酶的进展惊人。R18 能够复制长度为自身 7% 的序列,而最新的 tC9Y 核酶可以复制超过合成长度的 RNA 序列,至少在理论上,tC9Y 已经具备了自我复制的能力
在这一系列激动人心的科学发现中,克里克 1968 年的假说重新被人们翻了出来,而到了 1986 年,另一位诺贝尔奖得主、哈佛大学的沃特 • 吉尔伯特(Walter Gilbert)更是正式扛起了“RNA 世界”理论的大旗,要替 RNA 抢回地球生命的发明权了。
这可能是最接近真相也最能帮助我们理解生命起源的理论了。这个理论的核心就是,RNA 作为一种既能够存储遗传信息又可以实现催化功能的生物大分子,是屹立于生命诞生之前的指路明灯。可能在数十亿年前的原始海洋里,不知道是由于高达数百摄氏度的深海水温、刺破长空的闪电,还是海底火山喷发出的高浓度化学物质,数不清的 RNA 分子就这样被没有缘由地生产出来,飘散,分解。直到有一天,在这无数的 RNA 分子(也就是无穷无尽的碱基序列组合)中,有这样一种组合,恰好产生了自我复制的催化能力。于是它苏醒了,活动了,无数的“后代”被制造出来了。这种自我复制的化学反应所产生的大概还不能被叫作生命,因为它仍然需要外来的能量来源,它还没有“以负熵为生”的高超本领。但是,它很可能照亮了生命诞生前最后的黑夜,在它的光芒沐浴下,生命马上要发出第一声高亢的啼鸣。
今天,绝大多数地球生命都在中心法则的支配下生存繁衍,DNA 存储遗传信息并持续地自我复制,DNA 通过 RNA 控制了蛋白质的合成和生物体的性状(见图 3-8)。RNA 仅仅作为信息流动的中间载体出现,看起来多余而浪费。但是在远古地球,情形可能会很不相同。赞成 RNA 世界理论的科学家认为,在那时也许既没有 DNA 也没有蛋白质,而是 RNA 分子身兼两职:既能代替 DNA 存储遗传信息,又能代替蛋白质推动各种生物化学反应。
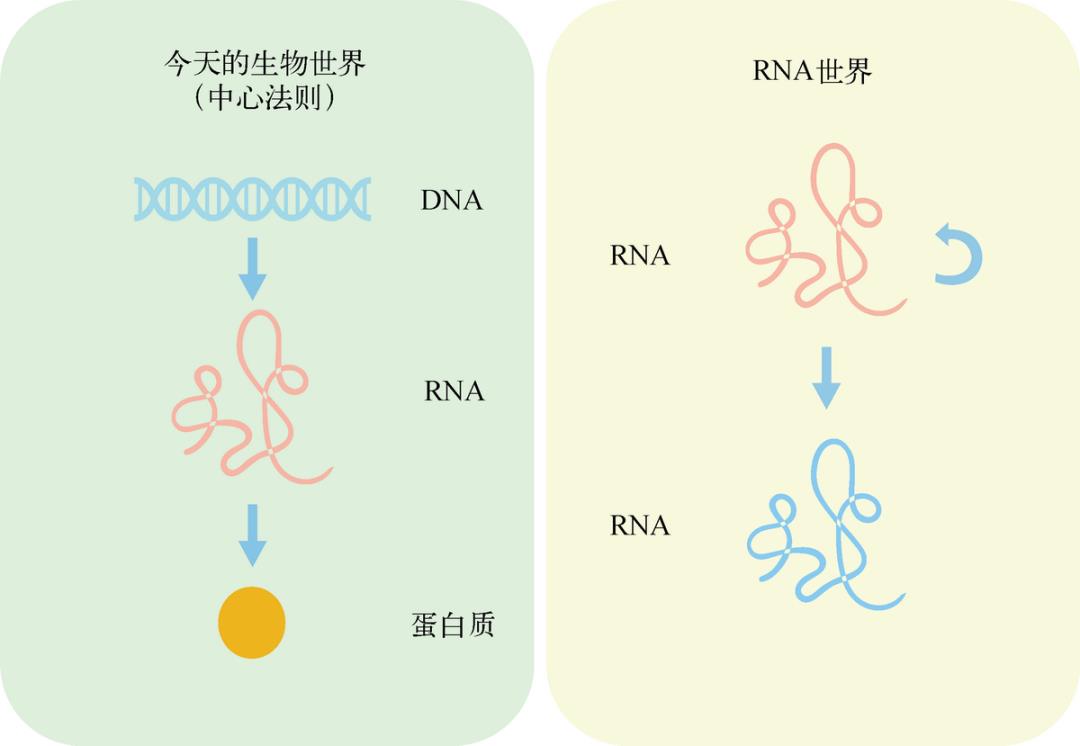
图 3-8 今天的生物世界和 RNA 世界
而可能在 RNA 世界出现甚至统治地球之后,今天地球生命的绝对统治者——DNA 和蛋白质——才崭露头角。从某种意义上说,它们利用各自的优势,从身兼存储遗传信息和催化生命活动的 RNA 分子那里抢走了原本属于它的荣耀。
相比 RNA,DNA 是更好的遗传信息载体,因为 DNA 的化学性质更加稳定,在自我复制的过程中出错率更低。相比 RNA,蛋白质又是更好的生命活动催化剂。由 20 种氨基酸装配而成的蛋白质分子,比仅有四种核苷酸装配而成的 RNA,可以折叠出更复杂的三维立体结构,可以推动更多更复杂的生物化学反应。因此,在今天的地球上,对于绝大多数生物而言,RNA 反而成了一个中间角色,仅仅通过生硬地将 DNA 插入蛋白质中的信息流动,宣示着自己曾经的无限荣光。仅仅在少数病毒体内,RNA 仍然扮演着独一无二的遗传信息密码本的角色。而我们推测,一些病毒之所以至今仍然顽固地抗拒使用 DNA 作为遗传物质,一个可能的原因是它们需要快速产生变异以逃脱免疫系统的攻击。在这方面,复制环节错误率较高的 RNA 分子反而具备了独特的优势。
所以说到这里,我们不得不遗憾地宣称,生命 1.0 到 4.0 的思想实验很可能是多余的。地球生命在诞生之初可能根本不需要独立的遗传物质和催化分子,它们只需要能量帮助地球
生命摆脱热力学的诅咒,在混乱无序的自然界中建立起精致有序的生命结构。而 RNA 祖先则肩负起自我复制、为生命开枝散叶的重任。而此后 40 亿年的漫漫演化,DNA 和蛋白质出现,多细胞生物诞生,人类萌生,智慧出现,其实都只是那一次伟大结合的绵绵余韵而已!
推荐阅读

作者:王立铭
* 浙江大学教授、科普作家、“文津图书奖”“吴大猷金奖”“菠萝科学奖”““全球华语科幻星云奖 ”得主王立铭科普力作
* 生命科学家饶毅、科幻作家刘慈欣、科幻作家郝景芳、资深媒体人罗振宇、德国癌症研究中心终身研究员刘海坤 联袂推荐
* 全景式解读地球生命的起源与演化,揭秘构成地球生命的十大要素,一本书带你全面了解生命科学
*回望40亿年演化历史,破解生命和智慧的奥秘,掌握和改变人类的命运


以上是关于DNA双螺旋结构发现70周年:自我复制的生命演化史!的主要内容,如果未能解决你的问题,请参考以下文章